

Macroecology deals with ecological patterns (e.g., species-area relationships – the increase of species richness with increasing area; 1-6; Fig SAR), and the underlying processes at large spatial scales. Although temporal changes in macrecological patterns, such as species geographic range and abundance distributions, have been apparent for more than a century and have frequently been quantified (e.g., 7-12), these changes remain largely unexplained phenomena of natural history, with an astonishing lack of consensus about the human role in these changes. Recent work has focused primarily on ecological explanations for these phenomena (e.g., 13,14, for review see 15). Although such analyses have provided important insights into the relationships between diversity, environmental variables and human activities, they reveal little about human impact on the evolutionary dynamics of ecosystems. The reason is that recent studies only compare sites along environmental gradients, and different sites may be driven by various forces caused by historical events (16). In conclusion, one of the reasons for the lack of consensus has been the absence of data and methods that can uncover the long-term dimension of human impact on the environment. More data and new methods, however, have become available during the last few years. As demonstrated by our unpublished analyses, these data shows changes that corresponds with for example the introduction of agriculture and migration periods. Unfortunatelly, we cannot analyse the consequences of the previous history without detailed archeological data for each focal site. These data can, however, be gathered from literature, regional databases, and museums.
Currently, various macroecological tools that can uncover changes in the forces that govern ecosystem dynamics have been developed (e.g., 17-28,5,6). These tools implicitly assume that the changes in the processes that govern the ecosystem dynamics are mirrored by changes in macroecological patterns. Therefore, the observable changes in macroecological patterns can reveal changes in the underlying processes.
The newest tools for macroecological analyses are designed to extract macroecological patterns from data from small plots, which is often the format of Palaeoecological and Archaeological data. Unfortunatelly not all the tools are available in the form of programme packages so far. As our team is capable of producing its own progamme packages, the intended project is the logical step forward.
For more information or opportunity to collaborate, please, contact sizling[at]cts.cuni.cz.
Species-Area Relationship (SAR)
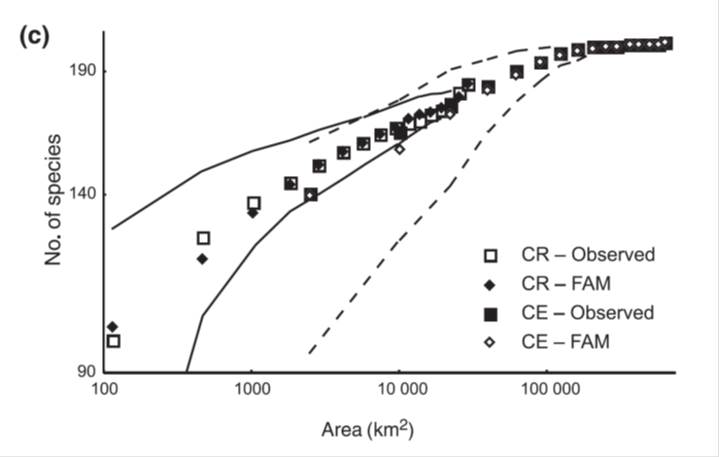
Species-Area Relationship (SAR) is one of the most important patterns studied by macroecologists. It links the area of a piece of landscape (or an island), and number of species that have their home in the piece of landscape (or the island). Standardly all SAR slows its increase as the area increases, showing an upper limit. This makes the rough pattern of SAR general. The rate of the increase is, however, taxon and biome specific, which makes the pattern unique in details.
The fig (adopted from 4) shows observed SARs within the Czech Republic and Central Europe (squares) and their 95% variations (full and dashed lines) as computed using bird atlas data. Diamonds show the predictions made by the finite area models (4). Macroecologists do both, computing patterns from rough data, and predicting the patterns from first principles and/or assumed underlying mechanisms.
References
- Tjørve E. (2003): Shapes and functions of species-area curves: a review of possible models. Journal of Biogeography 30:823-832.
- Tjørve E. (2009): Shapes and functions of species-area curves (II): a review of new models and parameterizations. Journal of Biogeography 36:1435-1445.
- Tjørve E., Tjørve K.M.C. (2012): Arrhenius and Gleason revisited: a new model approach resolves and old controversy. Journal of Biogeography 39:629-639.
- Šizling A. L., Storch D. (2004): Power-law species-area relationships and self-similar species distributions within finite areas. Ecology Letters 7:60-68.
- Šizling A. L., Kunin W., Šizlingová E., Reif J., Storch D. (2011): Between geometry and biology: the problem of universality of the species-area relationship. American Naturalist 175:602-609.
- Šizling A. L., Kunin W., Storch D. (2013): Taxon invariances, maximum entropy, and the species-area relationship: Reply to Harte et al. American Naturalist 181: 288-290.
- Gibson C. C., Ostrom E., Ahn T. K. (2000): The concept of scale and the human dimensions of global change: a survey. Ecological Economics 32:217–239.
- Stenseth N. C., Mysterud A., Ottersen G., Hurrell J. W., Chan K. S., Lima M. (2002): Ecological effects of climate fluctuations. Science 297:1292-1296.
- Walther G. R., Post E., Convey P., Menzel A., Parmesank C., Beebee T. J. C., Fromentin J. M., Hoegh-GuldbergI O., Bairlein F. (2002): Ecological responses to recent climate change. Nature 416:389-395.
- Meehl G. A., Washington W. M., Collins W. D., Arblaster J. M., Hu A., Buja L. J., Strand W. G., Teng H. (2005): How much more global warming and sea level rise? Science 307:1769-1772.
- Felde V. A., Kapfer J., Grytnes, J. A. (2012): Upward shift in elevational plant species ranges in Sikkilsdalen, central Norway. Ecography 35:922-932.
- Lenoir J., Virtanen R., Oksanen J., Oksanen L., Luoto M., Grytnes J.A., Svenning, J.C. (2012): Dispersal ability links to cross-scale species diversity patterns across the Eurasian Arctic tundra. Global Ecology and Biogeography 21:851-860.
- McKinney, M. L., and J. L. Lockwood. 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends in Ecology and Evolution 14:450–453.
- Reif J., Prylová K., Šizling A. L., Vermouzek Z., Šťastný K., Bejček V. (2013): Changes in bird community composition in the Czech Republic from 1982 to 2004: increasing biotic homogenization, impacts of warming climate, but no trend in species richness. Journal of Ornithology 154:359–370.
- Sutherland W.J. et al. (2006): The identification of 100 ecological questions of high policy relevance in the UK. Journal of Applied Ecology 43:617–627.
- Qian H., Ricklefs R. E. (2011): Latitude, tree species diversity and the metabolic theory of ecology. Global Ecology and Biogeography 20:362–365.
- Harte J., Kinzig A., Green J. L. (1999): Self-similarity in the distribution and abundance of species. Science 284: 334-336.
- McGill B. G., Etienne R. S., Gray J.S., et al. (2007): Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework. Ecology Letters 10:95–1015.
- Storch D., Šizling A. L., Reif J., Polechová J., Šizlingová E., Gaston K. J. (2008): The quest for a null model for macroecological patterns: geometry of species distributions at multiple spatial scales. Ecology Letters 11:771-784.
- Tjørve E. (2002): Habitat size and number in multihabitat landscapes: a model approach based on species-area curves. Ecography 25:17-24.
- Tjørve E. (2010): How to resolve the SLOSS debate: Lessons from species-diversity models. Journal of Theoretical Biology 264:604-612.
- Tjørve E., Kunin W.E., Polce C., Tjørve K.M.C. (2008): The species-area relationship: separating the effects of species abundance and spatial distribution. Journal of Ecology 96:1141-151.
- Tjørve E., Tjørve K.M.C. (2008): The species-area relationship, self similarity and the true meaning of the z-value; as revealed by converting z to species overlap. Ecology 89:3528-3533.
- Tjørve E., Turner W.R. (2009): The importance of samples and isolates for species-area relationships. Ecography 32:391-400.
- Turner W.R. Tjørve E. (2005): Scale-dependence in species-area relationships. Ecography 28:1-10.
- Šizling A. L., Šizlingová E., Storch D., Reif J., Gaston K. J. (2009a): Rarity, commonness and the contribution of individual species to species richness patterns. American Naturalist 174:82-93.
- Harte J., Smith A. B., Storch D. (2009): Biodiversity scales from plots to biomes with a universal species-area curve. Ecology Letters 12:789–797.
- Azaele S., Muneepeerakul R., Maritan A., Rinaldo A., Rodriguez-Iturbe I. (2009): Predicting spatial similarity of freshwater fish biodiversity. Proceedings of the National Academy of Sciences of the U.S.A. 106:7058–7062.